欢迎来到《实验技巧与小鼠百科》!本合集致力于成为实验台边最实用的伙伴,系统梳理那些让科研更高效、更规范的实验技巧,同时深入浅出地解读实验室小鼠背后的科学知识 。
在组织病理学研究中,Masson 染色(又称马松染色)作为经典的结缔组织染色方法,在区分胶原纤维与肌纤维、观察组织纤维化程度等方面具有不可替代的价值。
一、Masson 染色原理
Masson 染色本质上是一种多色复合染色法,其核心在于利用染料分子量与组织通透性差异,实现对不同纤维成分的选择性标记。
组织中的肌纤维、胶原纤维和细胞核等结构,在密度和理化性质上存在差异。染色过程中,小分子染料(如酸性品红、丽春红)能够较易渗入结构较为致密的肌纤维和细胞质,使其呈现红色;而大分子染料(如苯胺蓝、亮绿)则主要进入结构相对疏松的胶原纤维区域,赋予其蓝色或绿色色调。细胞核则通过苏木精染色显示为深蓝黑色,从而在镜下形成鲜明对比。
该技术特别适用于需要明确区分胶原纤维与肌纤维的病理场景,如评估肝纤维化程度、观察心肌修复后胶原沉积、鉴别肿瘤组织来源等,提供比常规 HE 染色更丰富的纤维结构信息。
二、Masson 染色实验准备与操作流程
试剂配制与保存
-
Weigert 铁苏木素染液
-
A 液(苏木素液):1g 苏木素溶解于 100ml 无水乙醇,室温避光保存。
-
B 液(铁媒染液):取 4ml 30%三氯化铁溶液,加入 95ml 蒸馏水与 1ml 盐酸,混匀后室温保存。
-
使用方法:临用前将 A 液与 B 液等体积混合,混合后不宜久存,建议当日使用完毕。
-
-
丽春红-酸性品红染液
-
称取丽春红 0.7g 与酸性品红 0.3g,溶于 99ml 蒸馏水中,充分搅拌溶解后,加入 1ml 冰醋酸,混匀。
-
过滤后使用,室温保存,若出现沉淀或颜色变化需重新配制。
-
-
1%磷钼酸水溶液
-
称取 1g 磷钼酸粉末,用少量蒸馏水溶解后,定容至 100ml。
-
室温保存,溶液澄清稳定。
-
-
苯胺蓝染液(或亮绿染液)
-
苯胺蓝液:取 2g 苯胺蓝溶解于 98ml 蒸馏水中,加入 2ml 冰醋酸,混匀后过滤使用。
-
亮绿液(替代选择):配制方法与苯胺蓝类似,可根据研究需要选择蓝色或绿色对比。
-
两种染液均需避光保存,长期放置后使用前建议重新过滤。
-
-
辅助试剂
-
0.5%醋酸水溶液:用于各染色步骤间的快速分化与清洗。
-
1%氨水或饱和碳酸锂溶液:用于苏木精染色后的返蓝处理。
-
样本前处理
-
固定:组织离体后应尽快置于足量 4%中性多聚甲醛固定液中,固定时间一般为 24-48 小时,确保完全穿透。避免使用含汞、铬的固定液(如 Bouin 液),以免干扰染色结果。
-
脱水、透明与包埋:固定后组织经梯度乙醇(70%、80%、95%、100%) 脱水,二甲苯或环保透明剂透明,最后浸入融化的石蜡中进行包埋。全过程需规范操作,避免组织收缩或硬化过度。
-
切片:使用切片机切取厚度为 4-5μm 的石蜡切片,裱贴于洁净或防脱载玻片上。切片应薄而均匀,无皱褶、无刀痕。60℃烘箱烤片 2-4 小时,使组织紧密贴附。
染色步骤简述
-
脱蜡与水化:彻底脱蜡是后续染色的基础,建议使用新鲜二甲苯与梯度乙醇。
-
核染色:Weigert 铁苏木素染色 5–10 分钟,流水返蓝,镜下控制至核清晰、背景干净。
-
浆染色:丽春红-酸性品红液染色 5–10 分钟,弱酸稍洗,去除浮色。
-
分化:1%磷钼酸处理至胶原区红色褪去、肌纤维仍红为止,此步需镜下实时监控。
-
胶原染色:直接入苯胺蓝或亮绿液染色 3–5 分钟,弱酸稍分化。
-
脱水封片:快速彻底脱水透明,中性树胶封固。
三、 Masson 染色关键环节与注意事项
-
分化步骤是整次染色的核心,时间不足则胶原着红色,过度则肌纤维红色变浅,必须镜下控制。
-
染色间隙切勿让切片干燥,否则易产生沉淀。
-
脱蜡与脱水透明不彻底会导致染色不均、背景模糊或后期褪色。
-
建议同步设置对照切片,便于结果判读与问题排查。
四、 Masson 染色常见问题分析与对策
染色偏红,蓝色过浅
- 可能由于磷钼酸分化时间不足,或苯胺蓝染色时间过短、染液失效。建议延长分化时间并镜下确认,检查并更换染液。
染色偏蓝,红色不明显
- 磷钼酸分化时间过长或丽春红-酸性品红染液失效、染色时间不足。建议缩短磷钼酸分化时间,确保丽春红染液新鲜有效,并可适当延长其染色时间。
核与背景模糊
- 多由苏木精染色过深、分化不足,或脱蜡不彻底导致。可缩短核染时间、加强分化,并确保脱蜡彻底。
切片出现杂质或沉淀
- 往往因为染色过程中切片局部干燥,或染液未过滤。应全程保持湿润,染液使用前过滤。
染色脱落
- 可能与组织本身状态、脱水透明不充分或切片过厚有关。建议使用防脱玻片,优化脱水流程,控制切片厚度。
五、 Masson 染色应用场景与总结
Masson 染色在病理诊断与研究中应用广泛,尤其适用于:
-
肝、肺、肾等器官纤维化程度的评估
-
心肌梗死后胶原修复的观察
-
某些肿瘤间质胶原分布的鉴别
-
皮肤、血管等结缔组织丰富部位的结构分析
一、H.E 染色技术原理与化学基础
H.E 染色(苏木精-伊红染色)是一种经典的组织学染色方法,其核心原理基于染料与细胞组分间特异性结合的化学特性,能够在光学显微镜下清晰区分细胞核与细胞质。
苏木精为碱性染料,其有效成分苏木红与媒染剂(通常为铝离子)结合形成带正电荷的色淀。该色淀与细胞核内带负电荷的脱氧核糖核酸(DNA)和核糖核酸(RNA)通过静电作用结合,将细胞核染成蓝紫色。
伊红为酸性染料,在适当 pH 条件下解离为阴离子,与细胞质、细胞外基质及大多数胞内蛋白质(特别是富含碱性氨基酸的蛋白质区域)的阳离子基团结合,将其染成粉红色至红色。细胞核与细胞质间显著的染色对比,为观察细胞形态、组织结构及病理变化提供了基础。
二、H.E 染色标准化实验操作流程
标准化的 H.E 染色操作流程是确保染色结果一致性、可靠性与可重复性的关键。
组织前处理
- 固定:离体组织应立即置于足量(体积比为 10:1 至 20:1)的 10% 中性缓冲福尔马林固定液或 4% 多聚甲醛固定液中。固定时间根据组织大小和类型而定,通常为 6 至 48 小时,旨在终止细胞自溶、保存抗原性并维持原始形态。
- 脱水与透明:通过梯度乙醇(如 70%、80%、95%、100%)逐步置换组织内水分。随后使用二甲苯等透明剂置换乙醇,使组织透化,便于石蜡浸润。
- 浸蜡与包埋:将透明后的组织置于熔融石蜡(熔点通常为 56-58℃)中充分浸润,随后用石蜡包埋成块,以提供切片所需的机械支撑。
切片与贴片
使用切片机将蜡块连续切为厚度 3-5 微米的薄片。切片在温水中展平后,捞取并贴附于经多聚赖氨酸或 APES 处理的防脱载玻片上。置于 60-65℃ 烤片机上烘烤 1-2 小时,使蜡片牢固粘附。
染色程序
H.E 染色的标准化操作流程可以概括为以下六个核心步骤,每个步骤都有关键的控制要点:
1、脱蜡与水化
- 主要操作:将已贴附组织的切片依次放入二甲苯、梯度浓度乙醇(如 100%、95%……),最后浸入蒸馏水。
- 作用与时间:此过程旨在彻底去除包埋组织的石蜡,并使组织重新水化,便于后续水溶性染液作用。通常在每缸试剂中处理 5-10 分钟。
- 关键控制点:必须确保脱蜡完全,任何石蜡残留都会严重阻碍染色。
2、细胞核染色(苏木精染色)
- 主要操作:将切片浸入苏木精染液(如 Harris 或 Mayer 苏木精)。
- 作用与时间:使细胞核着色。常规染色时间为 5-10 分钟,具体需根据染液的新旧程度和实验室室温进行微调。
- 关键控制点:染色时间不足会导致细胞核着色浅淡;时间过长则会导致染色过深,甚至使背景也非特异性着色。
3、分化
- 主要操作:将切片快速浸入 1% 盐酸乙醇(通常用 70% 乙醇配制)中。
- 作用与时间:快速浸提数秒至数十秒。目的是去除细胞质等非核结构上多余的苏木精染色,最好在显微镜下观察控制,直至细胞核轮廓清晰、背景基本无色为止。
- 关键控制点:这是决定细胞核染色特异性的最关键步骤。分化过度会将应有的核染色也洗脱,导致核染色丢失。
4、返蓝
- 主要操作:使用流动自来水长时间冲洗,或使用稀氨水、Scott 液等弱碱性溶液短时浸泡。
- 作用与时间:流水冲洗需 15 分钟以上,弱碱液浸泡约 30 秒至 1 分钟。目的是中和分化步骤留下的酸性环境,使苏木精在细胞核上的染色由红褐色恢复为稳定的鲜明蓝色。
- 关键控制点:返蓝不充分会导致细胞核呈现灰红色,严重影响核与细胞质之间的颜色对比度。
5、细胞质染色(伊红染色)
- 主要操作:将切片浸入伊红染液(常用浓度为 0.5%-1% 的水溶液或醇溶液)。
- 作用与时间:使细胞质、胶原纤维等结构着色。染色时间通常为 30 秒至 2 分钟。
- 关键控制点:染色时间过长容易导致细胞质着色过深,掩盖细微的形态结构。
6、脱水、透明与封片
- 主要操作:将染色后的切片依次通过梯度乙醇进行脱水,再经二甲苯透明,最后用中性树胶加盖玻片封固。
- 作用与时间:此步骤旨在永久保存染色结果。脱水需迅速,透明要彻底。
- 关键控制点:脱水不彻底会导致切片在进入二甲苯时出现浑浊;透明不彻底则会影响封片的透明度以及最终镜检的清晰度。
H.E 染色技术常见问题、原因排查与解决方案
一、染色过浅
主要可能原因
-
固定或脱水不彻底。
-
苏木精/伊红染色时间不足或染液失效。
-
分化过度。
解决方案与排查步骤
-
检查并优化前处理流程。
-
延长染色时间或更换新鲜染液。
-
严格控制分化时间,镜下监控。
二、染色过深
主要可能原因
-
苏木精染色时间过长。
-
分化不足或分化液失效。
-
返蓝不充分。
解决方案与排查步骤
-
缩短染色时间。
-
延长分化时间或更换分化液。
-
确保充分返蓝(流水或碱性液)。
三、核质对比度差浅
主要可能原因
-
分化不充分,背景过蓝。
-
返蓝不彻底,核色发灰。
-
伊红复染过深。
解决方案与排查步骤
-
精确分化至背景干净。
-
充分返蓝。
-
适当缩短伊红染色时间。
四切片脱落
主要可能原因
-
烤片时间或温度不足。
-
载玻片未进行防脱处理。
-
组织脱水、透明不彻底。
解决方案与排查步骤
-
确保充分烤片(60-65℃,≥1小时)。
-
使用防脱载玻片。
-
检查并优化脱水、透明步骤。
五、非特异性沉淀
主要可能原因
-
染液有沉淀或杂质。
-
脱水用乙醇或二甲苯不纯。
-
切片干燥。
解决方案与排查步骤
-
定期过滤染液。
-
使用优质纯试剂。
-
染色过程中保持切片湿润。
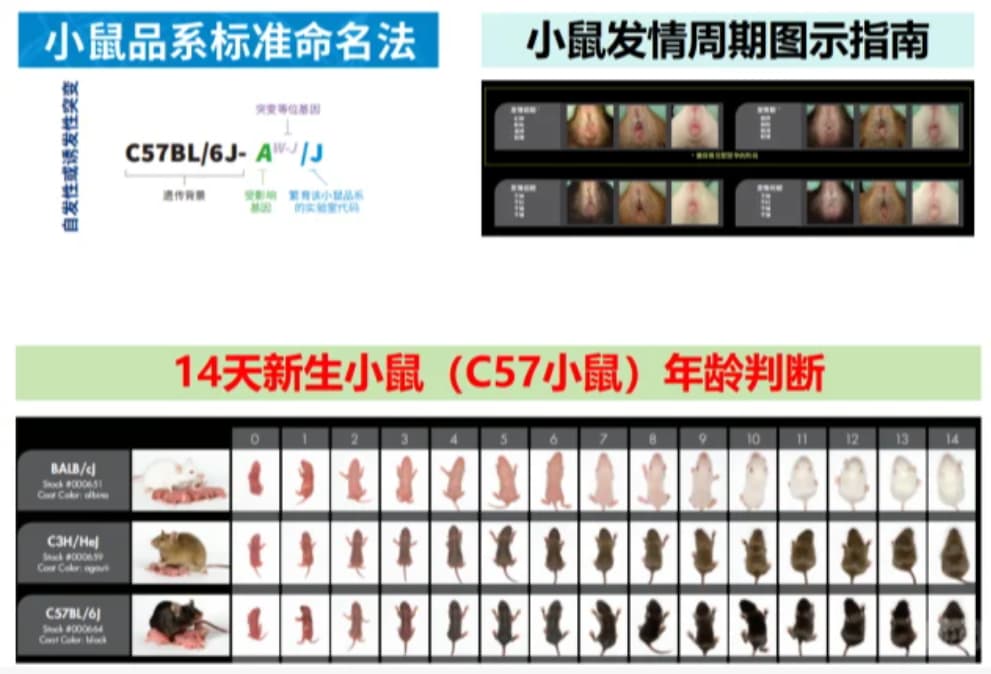
刚养 C57 小鼠总踩坑?14 天新生鼠分不清、发情周期看不准、品系名看不懂?这篇精简版笔记直接抄作业!
养 C57 小鼠必看!14 天新生鼠年龄判断 + 发情周期 + 品系命名,一篇搞定
14 天新生 C57 鼠年龄判断(肉眼秒会)
体重:约 4-6g(掂量有轻微坠手感)
外观:毛长 1-2mm,能看清背部毛发纹理,耳朵完全立起不贴脸
行为:会缓慢爬动,能短距离追着母鼠跑,不会跳
发情周期 4 阶段(附简易判断法)
发情前期
发情期
发情后期
间情期
小鼠品系命名法(C57 相关必懂)
C57BL/6J 为例:
C57:近交系核心编号
BL:毛色(Black,黑色)
6:亚系编号
J:来源机构(Jackson 实验室)
→ 简单说:C57 背景 + 黑色毛 + 6 号亚系 + J 实验
![]()
在免疫学、传染病、癌症生物学、造血干细胞和再生医学等领域的研究中,免疫缺陷小鼠已经成为十分重要的模型工具。什么是免疫缺陷小鼠?它们是怎么来的?目前免疫缺陷小鼠的品系繁多,如何选择合适的免疫缺陷小鼠模型呢?
免疫缺陷小鼠大盘点!内容超级详细超级丰富!

免疫缺陷小鼠有哪些品系?特性分别是什么?
一、无胸腺裸鼠
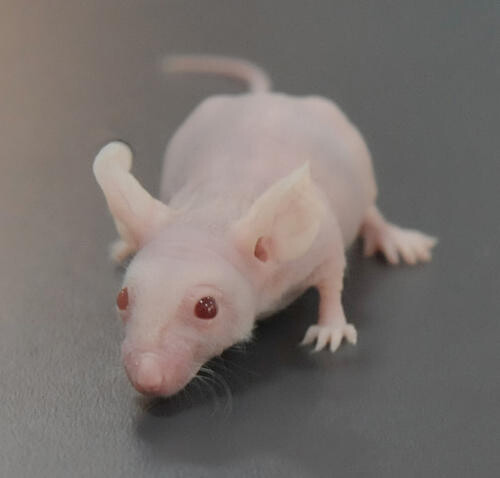
1962年,在英国格拉斯哥Ruchill医院的病毒实验室中,病毒流行病学家Norman Grist“偶遇了”一只全身无毛的小鼠,这就是第一只被发现的裸鼠。两年后,爱丁堡动物遗传研究所的S.P. Flanagan证实这是不同与一般无毛小鼠的突变种,取名为nude小鼠。
裸鼠的主要特征表现为无毛(Hairless)和无胸腺(Athymus)。这些表型归咎于Foxn1蛋白的隐形突变。无毛的成因并不是毛囊缺失,而是由于Foxn1功能缺失导致毛发无法穿透表皮,盘绕在基底层内。
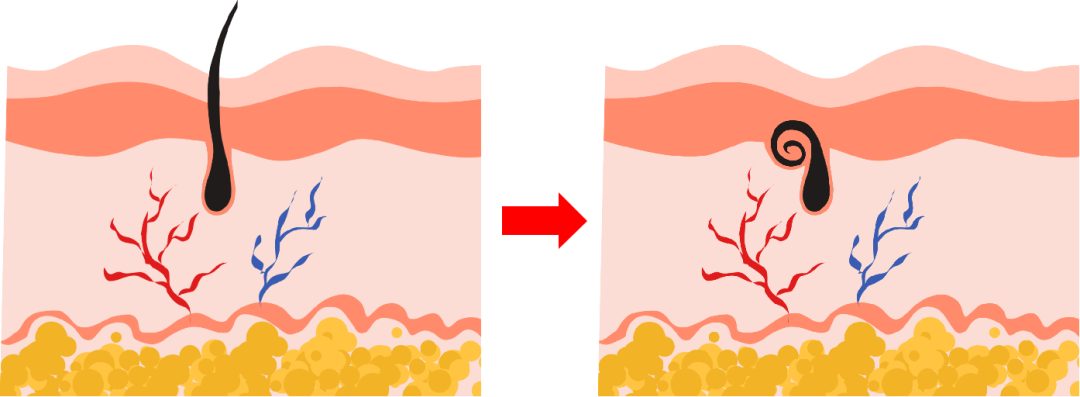
小鼠无胸腺同样是由于Foxn1功能缺失,导致胸腺上皮细胞和T细胞祖细胞无法正常增殖和分化,影响胸腺的发育和成熟。由于辅助T细胞的缺失,B细胞的发育也受到影响。
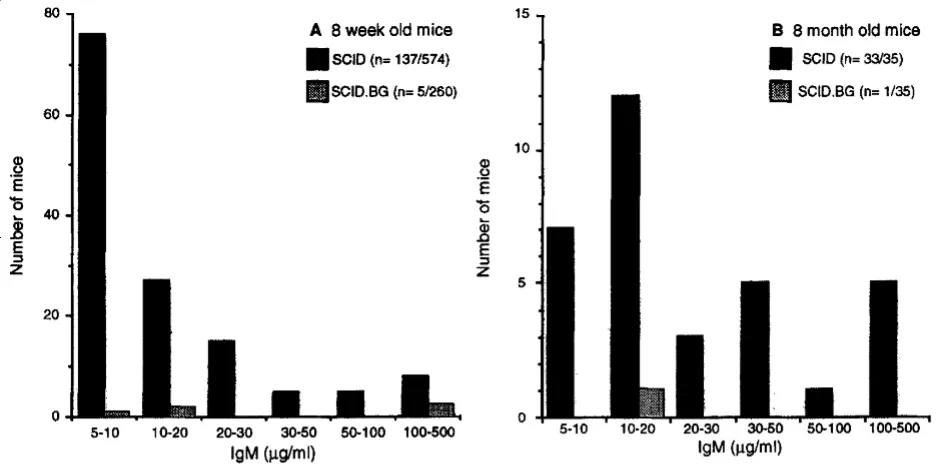
裸鼠先天无胸腺,生长发育不良,繁殖力低下;易发生严重感染,抗体主要为IgM,只有少量IgG无接触敏感性,无移植排斥反应,因此是非常好的肿瘤研究模型,被广泛用于人类肿瘤、免疫缺陷疾病和疾病发生机理等方面的研究。裸鼠能够移植人源或鼠源的肿瘤细胞系,且由于无毛更易评估皮下成瘤的生长和活体成像观察。但是,裸鼠的NK细胞活性增强,体液免疫能力正常,免疫缺陷程度仍较低,不适于进行患者来源肿瘤的异种移植(patient-derived xenografts, PDX)。
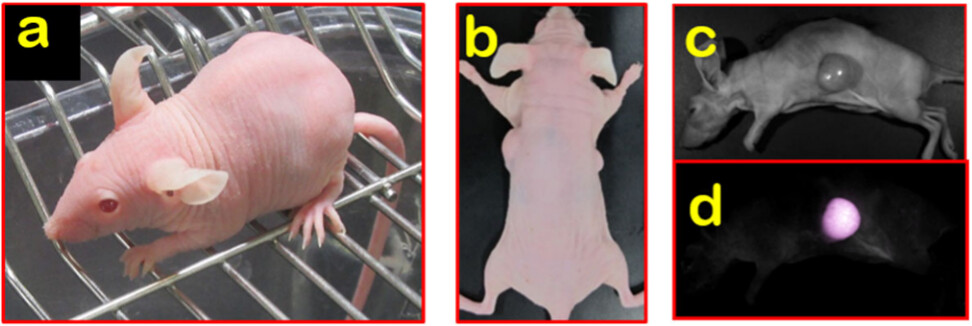
图3 (a) BALB/c Nude Rag-2/Jak3双缺陷(Nude R/J)无毛表型;(b) 直接观察Nude R/J的皮下肿瘤结节;(c-d) 观察Nude R/J的荧光信号。
Okada S, Vaeteewoottacharn K, Kariya R. Application of Highly Immunocompromised Mice for the Establishment of Patient-Derived Xenograft (PDX) Models [J]. Cells, 2019, 8.
Foxn1基因位于第11对染色体上,常用“nu”表示裸基因符号。将裸基因“nu”导入其他品系小鼠中可获得不同的突变系。常用的裸小鼠突变系有BALB/c–nu、NC-nu、C3H-nu、Swiss-nu等。
由于裸鼠的乳腺发育不完全,裸鼠的繁育方式主要是通过雌性杂合子和雄性纯合子进行交配。
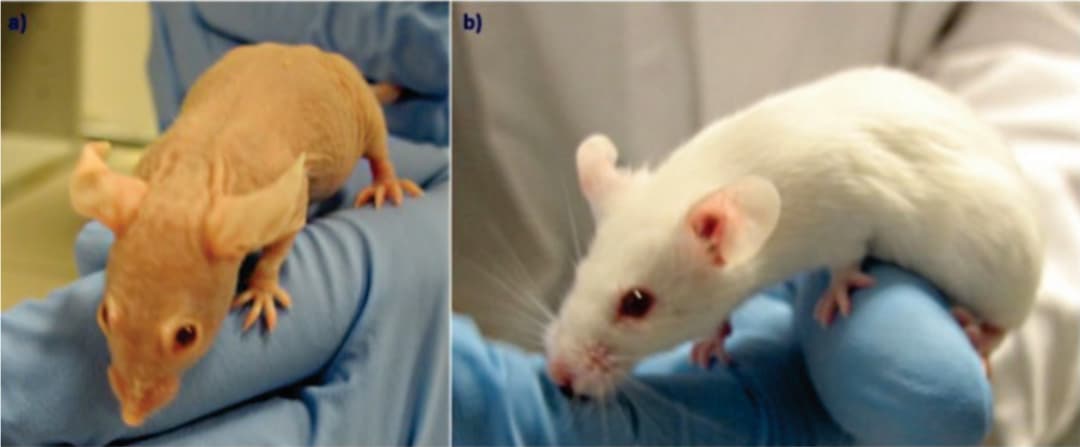
图4 a:雌性裸鼠纯合子(nu/nu),b:雌性裸鼠杂合子(nu/+)
Szadvari I, Krizanova O, Babula P. Athymic nude mice as an experimental model for cancer treatment. Physiol Res. 2016;65(Suppl 4):S441-S453. doi:10.33549/physiolres.933526
二、CBA/N小鼠

CBA/N小鼠1966年起源于CBA/H品系,1972年证实为X-链隐性突变系,命名为X-链免疫缺陷型,基因符号为xid,位于X性染色体上。CBA/N小鼠B细胞功能缺陷,对T细胞非依赖性II型抗原(如聚蔗糖、右旋糖苷、肺炎球菌多糖体)不能引起应答反应,而对T细胞非依赖性I型抗原(如:布氏菌脂多糖等)呈正常反应;分泌IgM和IgG亚类的B细胞数量减少,其T细胞功能正常。

三、Scid小鼠

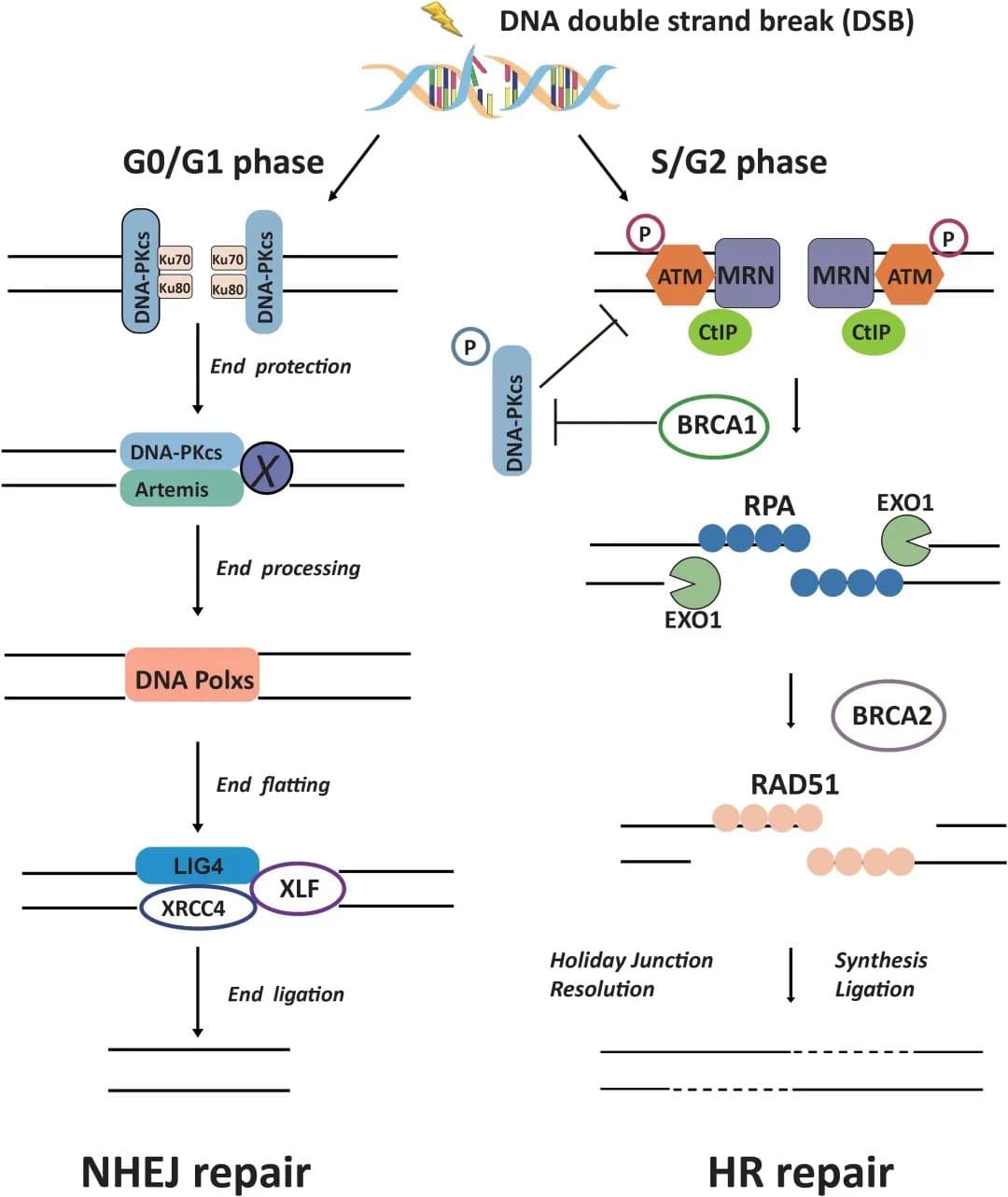
SCID是由Severe Combined Immune-deficiency缩写而来,这一症状最早是1950年在人类的婴儿中发现,1983年美国Bomsa在CB17近交系小鼠中发现位于16号染色体上的Prkdc基因存在隐性突变,并首次描述了缺乏功能性T和B淋巴细胞的严重联合免疫缺陷(Severe Combined Immunodeficient Mice,SCID)小鼠。
Scid小鼠外观与普通小鼠差别不大,有毛,被毛白色,体重发育正常。免疫细胞发育分化环境正常,但胸腺、脾脏、淋巴结等组织器官中无功能性T细胞及B细胞(重量不及正常的30%),组织学上表现为淋巴细胞显著缺陷。Prkdc基因编码了DNA依赖性蛋白激酶的催化亚基(DNA-PKcs)。该蛋白是DNA修复中NHEJ通路的关键蛋白,并且参与了T、B 细胞的T细胞抗原受体(TCR)、B 细胞抗原受体(BCR)基因的V(D)J重排。SCID小鼠Prkdc的缺失导致TCR、BCR基因的V(D)J重排无法进行,进而影响T、B细胞的成熟。

Scid小鼠胸腺多位脂肪组织包围,没有皮质结构,仅残存髓质,主要由类上皮细胞合成纤维细胞构成,边缘偶见灶状淋巴细胞群。脾白髓不明显,红髓正常,脾小体无淋巴细胞聚集,主要由网状细胞构成。淋巴结无明显皮质区,麸皮质区缺失,由网状细胞占据。小肠粘膜下和支气管淋巴集结较少见,结构内无淋巴聚集。

四、Beige小鼠
Beige小鼠为NK细胞活性缺陷的突变系小鼠,在第13号染色体上的隐性遗传基因bg发生突变引起,这种动物被称为Beige (bg/bg)小鼠。
Beige小鼠内源性NK细胞发育和功能缺陷,血液凝固和巨噬细胞活性也有缺陷,其免疫抗肿瘤杀伤作用出现较晚,缺乏细胞毒性T细胞功能,对同种、异种肿瘤细胞的体液免疫功能减弱,欠缺巨噬细胞的抗肿瘤活性、杀伤细胞活性等。Beige小鼠常应用于免疫学领域,对各种致病因子较敏感,需良好SPF环境。

五、SCID Beige小鼠
SCID Beige小鼠是在20世纪90年代通过将Beige突变引入到CB17 SCID小鼠中而形成的。SCID Beige小鼠在功能性T、B细胞缺失的基础上,新引入Beige突变使NK细胞的活性也受损,进而使免疫缺陷程度进一步提高。同时,SCID Beige小鼠的免疫渗漏率小于2%且不随年龄增长而增加。因此,与C.B-17 SCID小鼠相比,SCID Beige小鼠上某些细胞系或组织的移植成瘤率较高。此外,SCID Beige小鼠的先天性免疫细胞功能仍旧正常,因而可被用来模拟并评估CAR-T疗法引发的细胞因子释放综合症。
六、RAG-/-小鼠

1992年,缺失重组激活基因(Rag-/-)的免疫缺陷小鼠模型问世。重组激活基因包括Rag1和Rag2 ,两者编码的RAG1和RAG2组成的RAG复合体在TCR和免疫球蛋白V(D)J重排中起到关键作用。RAG复合体识别RSS序列并将DNA双链剪切,之后DNA双链通过DNA-PKcs、Ku70/Ku80、Artemis等酶启动NHEJ,修复DNA双链完成VDJ重排。因此,Rag1和Rag2任一基因的缺失都会导致T细胞和B细胞的缺乏。
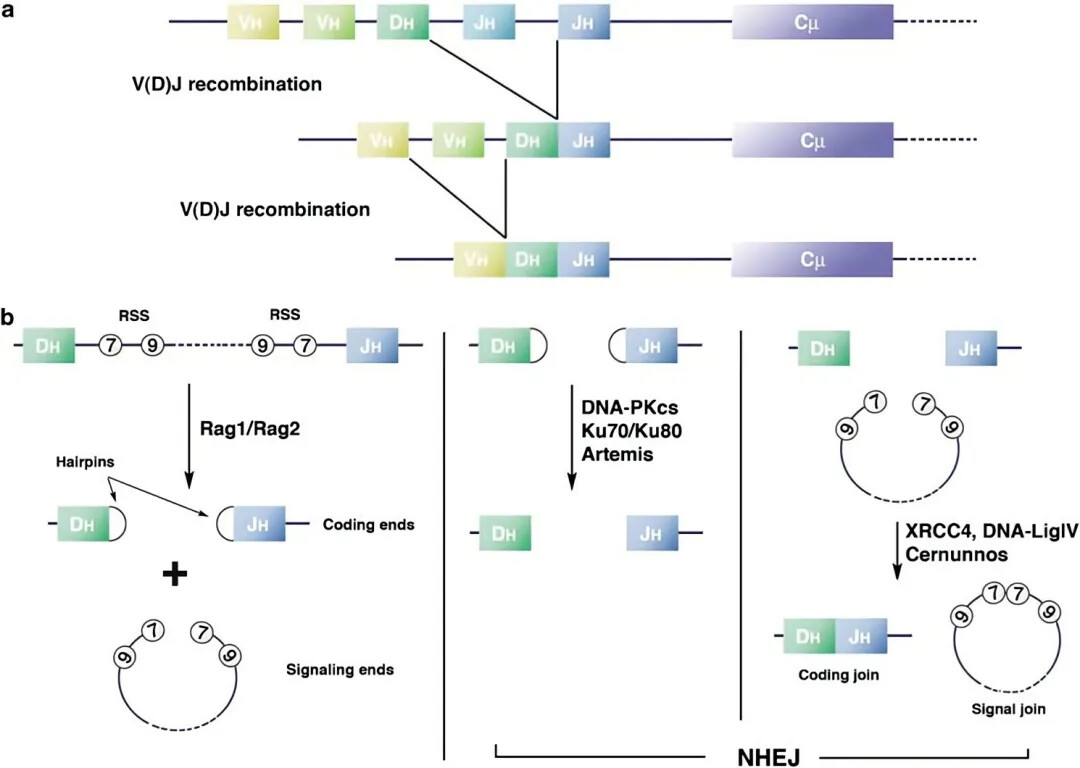
Rag1-/-和Rag2-/-小鼠的特征基本相似。与SCID小鼠相比,Rag基因缺失能更早且更完全地阻断T和B细胞分化。并且Rag基因缺失不影响DNA双链修复进程,因此Rag-/-小鼠对辐照不敏感。不过,Rag-/-小鼠保留了较高水平的NK细胞,在一定程度上也限制了人HSC细胞的移植。主要用于造血和免疫系统缺陷中的Rag基因功能研究,癌症和毒理研究(作为移植宿主)以及血液学,免疫学和炎性的研究。

七、NOD SCID小鼠

1980年, Makino(日本Shionogi公司)发现非肥胖糖尿病(NOD)小鼠患有由T淋巴细胞浸润和胰岛β细胞破坏引起的糖尿病,同时还伴有多种免疫异常,包括补体缺失和NK、巨噬细胞和树突状细胞功能受损等。
NOD SCID小鼠诞生于1995年,是在SCID小鼠的基础上,通过与非肥胖性糖尿病小鼠NOD/Lt杂交而成的一种重度免疫缺陷动物。NOD小鼠本身就存在一系列先天性的免疫细胞功能受损,包括NK细胞功能不足,缺乏循环补体,以及抗原递呈细胞分化和功能的缺陷。同时,在NOD小鼠中,巨噬细胞上的信号调节蛋白-α(SIRPα)抑制性受体与人源细胞的CD47间具有更大的结合亲和力,因而可以有效地阻止人源移植细胞被巨噬细胞吞噬。因此,引入了SCID基因突变的NOD小鼠就兼具T、B细胞缺失、NK细胞功能受损、补体结合能力减弱、抗原递呈细胞的免疫调节功能受损、巨噬细胞对人源细胞清除减少的特点;各种肿瘤细胞可以植入,且较少发生排斥反应及移植物抗宿主病(GVHD)。其免疫缺陷程度高但是免疫渗漏少,一度成为应用最广的免疫缺陷模型。NOD SCID小鼠改善了人源外周血单核细胞(PBMCs)和造血干细胞(HSCs)的移植成功率,且可进行患者来源的异种移植(PDX)。但是,NOD SCID小鼠的胸腺淋巴瘤发生率较高,在无特定病原体条件下,其平均寿命只有8.5个月;用于肿瘤、免疫学、胚胎干细胞、艾滋病及肝炎研究,为T、B、NK细胞缺陷的严重联合免疫缺陷动物模型。
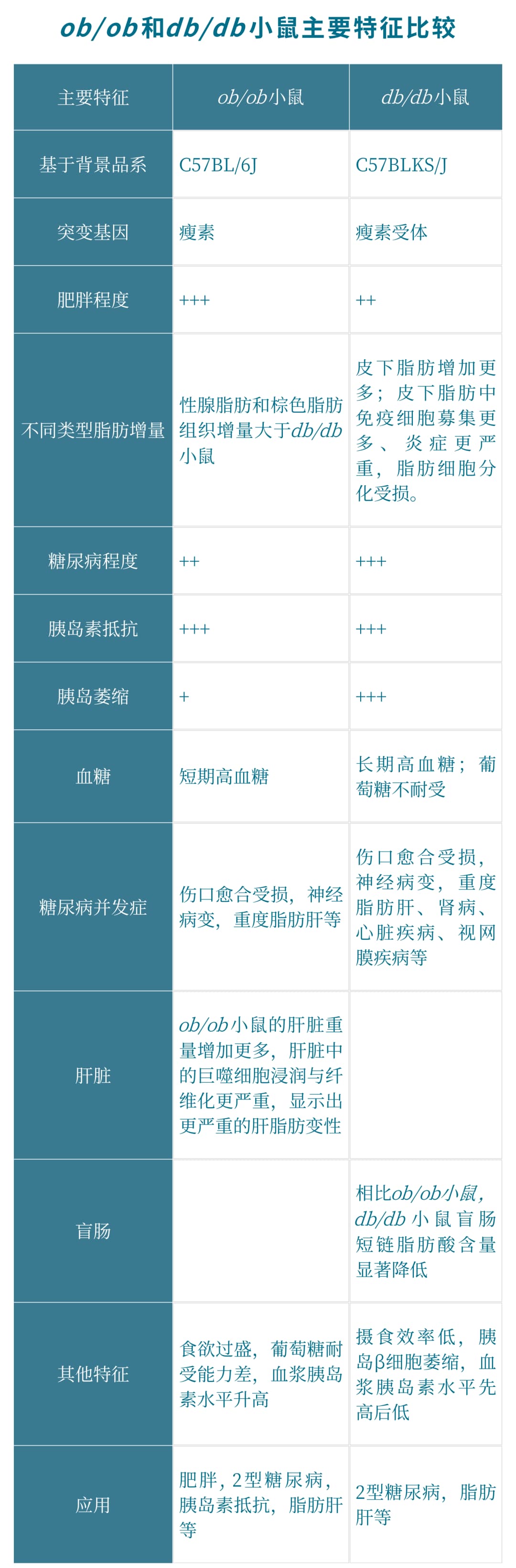
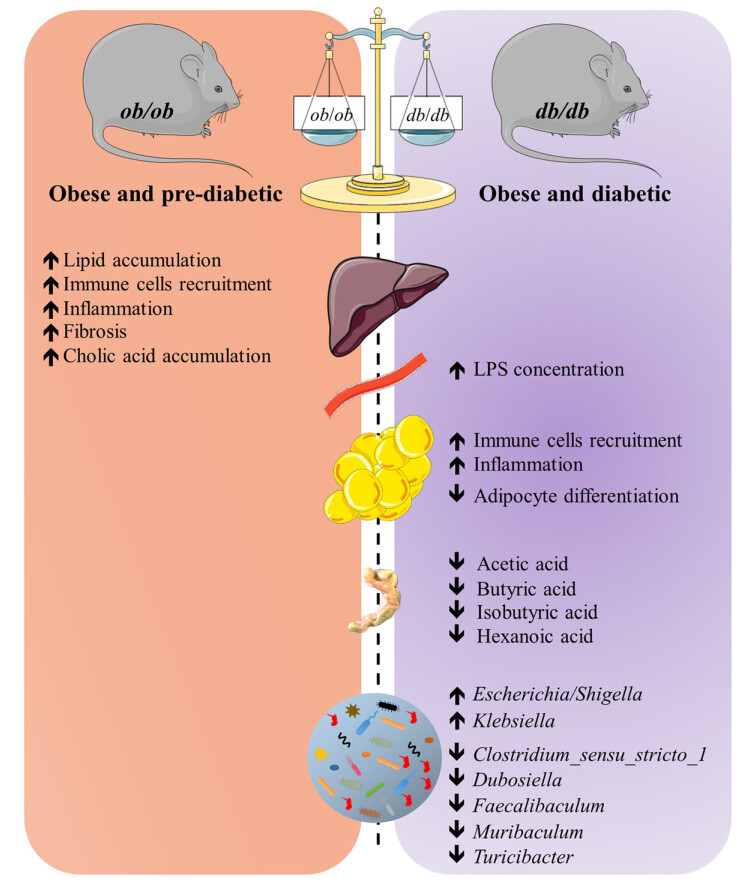
突变系小鼠ob/ob 和db/db 作为肥胖症与糖尿病的研究模型在代谢相关研究中广泛应用。它们为什么起名ob/ob 和db/db ?它们是怎么被发现的?又有什么差异呢?
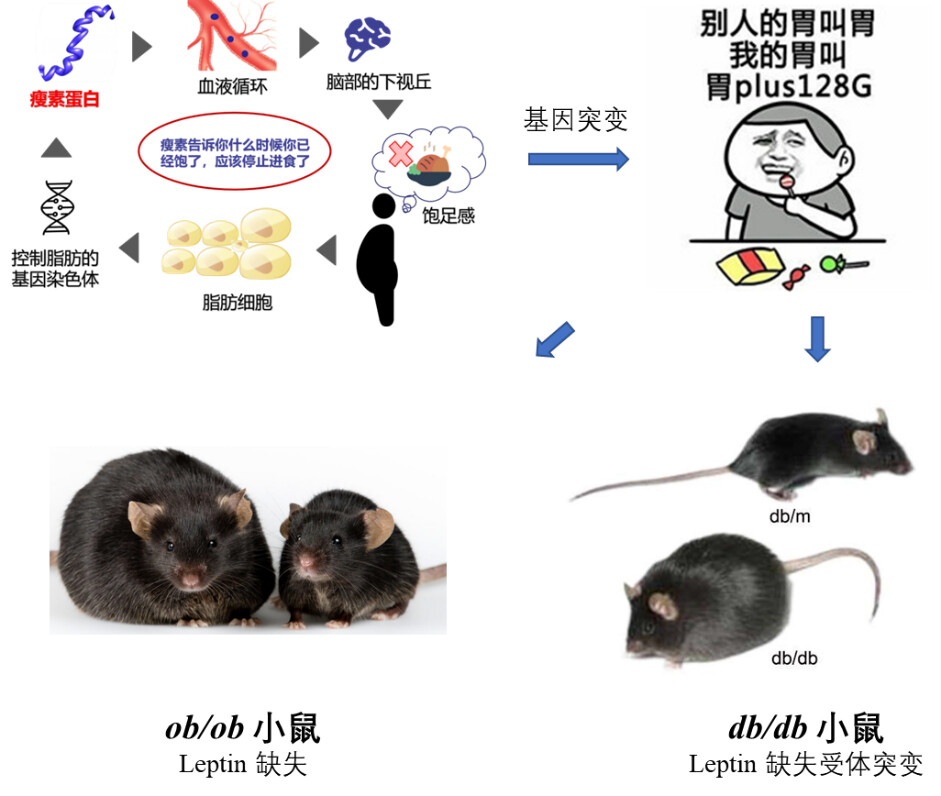
ob/ob 和db/db 小鼠分别最早于1949年和1969年在Jackson实验室被Joseph Coriell和Douglas Coleman研究发现。它们都携带隐性突变,并且都会产生明显的肥胖,其中一种小鼠表现出严重肥胖和轻度糖尿病,而另一种小鼠出现严重糖尿病并同样表现出肥胖特征。它们的名字也就由此而来,表现为严重肥胖的鼠命名为ob/ob小鼠 (Obesity,简写为ob ),表现为严重糖尿病的小鼠命名为db/db 小鼠(Diabetes,简写为db )。

什么是瘦素?
听到瘦素,大家可能都会想,瘦素是不是一种和变瘦有关的物质啊?
其实确实如此。**瘦素是由一条肽链构成的肽类激素。**主要由脂肪细胞分泌,其表达主要在白色脂肪组织,同时在心肌、骨骼肌、胎盘、肺、乳腺上皮和胃黏膜等位置均有表达。分泌到血液中的瘦素会穿过血脑屏障进入大脑,向中枢神经系统传达胃肠饱胀信号。我们因进食过多,脂肪增加时,过多的脂肪就会产生瘦素,进而刺激下丘脑的感受器,通过交感神经将超重信号传递到摄食中枢,大脑会“命令”我们减少摄食,增加能量消耗,以保持体重的恒定。然而,当我们处于饥饿状态时,血液中的瘦素水平下降,会进而刺激下丘脑提高副交感神经的兴奋性,增加食欲,降低能量消耗,以恢复正常体重。所以瘦素通过调节食欲来帮助我们维持体重稳态。瘦素(Lep)或瘦素受体(Lepr)的突变会导致饮食失调,这时瘦素信号不起作用,过高的食欲增加脂肪累积,进一步诱发肥胖、胰岛素抵抗、脂肪肝及糖尿病等多种代谢性疾病。
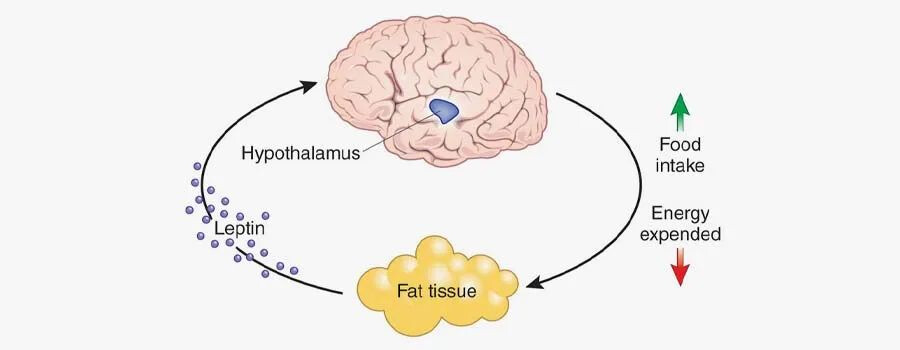

我们常用于肥胖症相关研究的ob/ob和db/db小鼠模型就是基于瘦素或者瘦素受体基因构建的。简言之:
ob/ob 小鼠瘦素合成分泌障碍,瘦素受体正常,突变位于6号染色体。
db/db 小鼠瘦素合成分泌正常,瘦素受体缺陷,突变位于4号染色体。
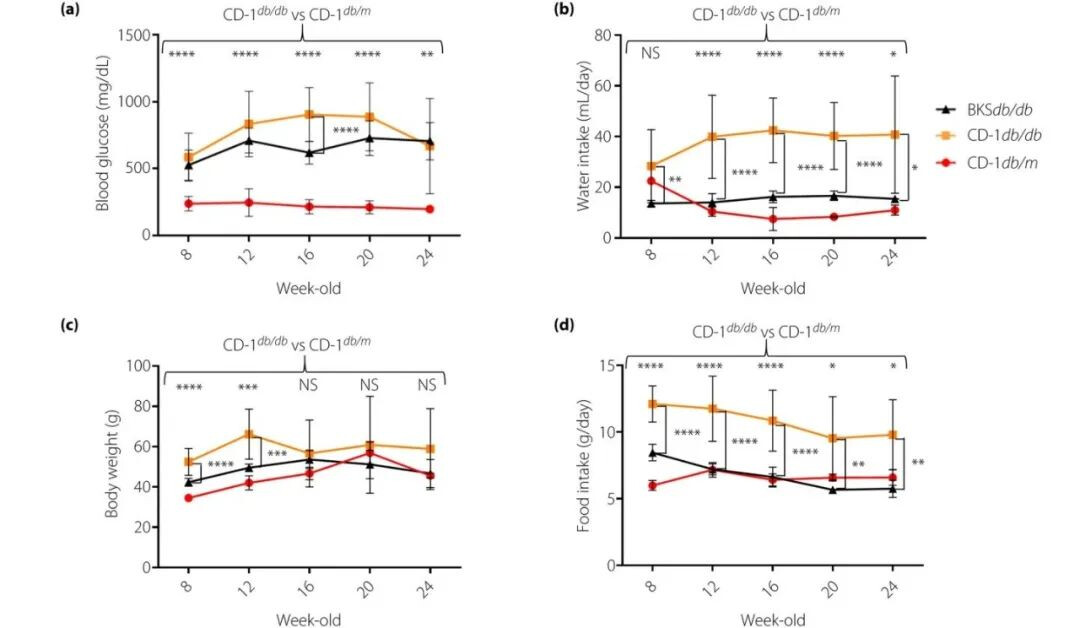
一、db/db 小鼠
db/db小鼠是Leptin受体点突变导致leptin信号通路障碍,为糖尿病易感性,出现重度糖尿病症状,胰岛β细胞严重受损,仅存活约10个月。
主要特征:db/db小鼠出生后6周即可出现明显的肥胖和空腹血糖增加,饮水量、尿量增加,8-12周时最明显,并可出现糖尿病肾病等并发症。小鼠表现出肥胖、胰岛素抵抗、高血糖、脂肪肝等症状。
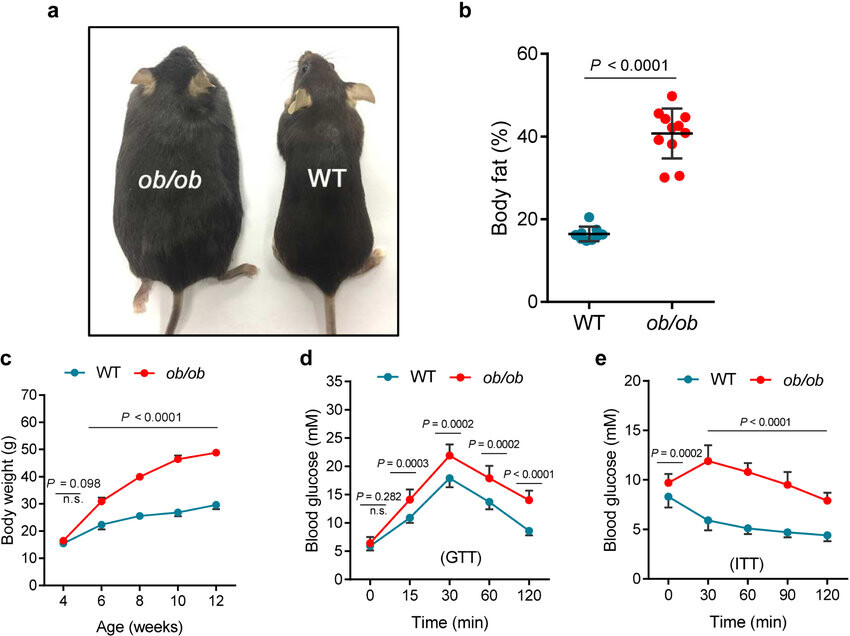
二、ob/ob 小鼠
ob/ob小鼠是瘦素基因(ob)纯合突变的小鼠,其特征为肥胖、多食、高血糖和胰岛素抵抗。
主要特征:在2周龄时肥胖个体外表上有别于正常个体。在8~9月龄时,动物体重增加到约为70克,多种代谢失调,包括脂肪形成增加,脂肪分解减少。代谢失调与过食症和胰岛素分泌过多有关,动物在6~9周龄时有中度的高血糖,但12~16周后自发消失。
远交群(outbredstock),又称封闭群(closedcolony),指的是在某一群体内,以非近亲交配方式培育而成的小鼠。远交群由于连续15代不从外部引入新种群,群体遗传背景较稳定,但个体差异较大,远交群环境适应力较强且抗病能力较强繁殖力强,价格较为经济,广泛用于药理学及毒理学研究。
一、KM小鼠

即昆明小鼠,白色,我国特有的封闭群小鼠,具有独特的遗传学性状,有别于NIH等国际知名的封闭群。一直是我国生产量、使用量最大的远交群小鼠,该鼠特点是繁殖率和成活率高,抗病力强,适应性强。广泛应用于药理、毒理、微生物学的研究以及药品、生物制品的检定。
昆明小鼠品系名称的由来:1926年美国Rockfeller研究所从瑞士引入白化小鼠培育成Swiss小鼠。1944年3月17日卫生部北京生物制品研究所汤飞凡从印度Hoffkine研究所引进Swiss小鼠,饲养在中国昆明中央防疫处。1952年由昆明引入北京生物制品所,1954年推广到各地。由于该小鼠起初引入地是昆明,故称之为昆明小鼠。

二、NIH小鼠
白色,由美国国立卫生研究院(NIH)培育而成。1980年引人北京生物制品研究所。该鼠适应性强,繁殖力强,产仔成活率高,雄性好斗,免疫反应效感性比昆明小鼠强。是国际上广泛通用的实验动物,已选为某些生物制品检定实验的法规动物。广泛用于药理和毒理研究以及生物制品检定。
三、ICR小鼠

白色,是国际通用的封闭群小鼠,为美国 Handshake研究所饲养的瑞士种小鼠。由美国肿瘤研究协会分送各地,取名为ICR。我国1973年从日本国立肿瘤所引进。该品系小鼠温驯,健壮,繁殖力强,适应性强,生长快,实验重复性较好,自发性肿瘤发生率低;广泛用于药理、毒理、肿瘤、放射性、食品、生物制品等的科研、生产和教学。
ICR小鼠名称的由来:ICR小鼠是由美国癌症研究所命名,ICR为(美国)癌症研究所(Institute of Cancer Research)的三个首写字母组成。后来被世界各地引进,成为国际通用的封闭群小鼠。
四、LACA小鼠
白色,1935年英国Carworth公司从美国Rockfeller研究所引进,经20代近交培育后,采用随机交配繁殖,后引入英国实验动物中心改名为LACA,我国1973年从英国实验动物中心引进。
五、CFW小鼠
白色,CFW小鼠是1973年由日本国立肿瘤研究所引入我国的。该小鼠起源于Webster小鼠,1935年英国Carwarth从Rockeffler研究所引进,经过20代近亲兄妹交配后,采用随机交配而成。
前面多次提到了瑞士种小鼠,这到底是什么品系呢?


一、BALB/c小鼠(白色)

特点:
(1)乳腺肿瘤自然发生率低(10%~20%),但用乳腺瘤病毒诱发时发病率高,卵巢、肾上腺和肺的肿瘤、白血病在该小鼠有一定的发生率。肺癌发病率雌性26%,雄性29%。白血病发病率雌性12%,雄性10%。血压与其他近交系小鼠相比为最高,有自发高血压症。
(2)易患慢性肺炎;对鼠伤寒沙门氏菌补体敏感,对麻疹病毒中度敏感,对利什曼原虫属、立克次氏体和百日咳组织胺易感因子敏感。
(3)对放射线甚为敏感;有自发高血压症,老年鼠心脏有病变,雌雄鼠均有动脉硬化。
(4)与其他近交系相比,肝、脾与体重的比值比较大,20月龄的雄鼠脾脏有淀粉样变。
(5)用途:广泛地应用于肿瘤学、生理学、免疫学、核医学研究,以及单克隆抗体的制备等。

二、C3H/He小鼠(野鼠色)

特点:
(1)乳腺癌发病率高,6~10月龄雌鼠乳腺癌自然发生率达85%-100%,乳腺癌通过乳汁而不是胎盘途径传播。
(2)补体活性高,干扰素产量低。
(3)对狂犬病毒敏感,对炭疽杆菌有抵抗力。
(4)血液中过氧化氢酶活性高。雄鼠对氨气、氯仿、松节油等甚为敏感,死亡率高。
(5)红细胞及白细胞数较少。
(6)仔鼠下痢症感染率高。
(7)对致肝癌因子敏感:14月龄雌性小鼠肝癌发率为85%。

三、C57BL/6小鼠(黑色)

特点:
(1)乳腺癌发病率低,对放射性物质耐受强,但照射后肝癌发病率增高。
(2)眼畸形,口唇裂的发生率达20%。
(3)淋巴细胞性白血病发病率为6%。
(4)对结核杆菌、百日咳组织胺易感因子敏感,对鼠痘病毒有一定抵抗力。
(5)嗜酒精性高,肾上腺素类脂质浓度低。

四、DBA/2小鼠(浅灰色)

特征:
(1)乳腺癌发生率经产雌鼠为66%,未产鼠为3%;雌雄鼠均可自发产生淋巴瘤;抗DBA/1所生长的瘤株,但黑色素瘤S-91在DBA/1和DBA/2小鼠中均能生长。
(2)白血病的发生率,DBA/2/Ola雌鼠为34%,雄鼠为18%,而DBA/2N雌鼠为6%,雄鼠为8%;雄鼠接触氯仿和乙二醇的氧化物,以及缺乏维生素K时,死亡率高;35日龄鼠听源性癫痫发作率为100%,55日龄后为5%。
(3)对鼠伤寒沙门氏菌C5有抵抗力,对百日咳组织胺易感因子和酒精过敏。
(4)老龄雄鼠有钙质沉着。

五、A系小鼠

特点:
(1)迟发性进展性的肌肉萎缩(进行性肌肉萎缩模型,dysferlinopathy模型)
这是由于该系小鼠含有天然发生的dysferlin突变,在dysferlin(Dysf)基因的4个内含子中插入了一个约5-6kb大小的ETn反转录转座子。肌肉萎缩首先是在腹部,然后是近端肌肉,再到远端肌肉,近端肌肉比远端肌肉严重。
(2)肾上腺皮质激素诱导先天性腭裂
(3)癌症模型
高发生率的自发肺部腺瘤。对致癌剂敏感,容易引起肿瘤的发展。多产雌性小鼠自发性高发生率乳腺腺癌。
(4)感染模型
用于感染和免疫学研究。A小鼠对猪链球菌具有高度反应性,特别是感染的急性败血症期的反应可以是6倍(100%对16%)于C57BL/6小鼠的反应。由于猪链球菌是猪和人类的病原菌,因此,A小鼠适用于研究猪链球菌感染的体内反应机制。
(5)睡眠相关的癫痫模型
研究发现,在杰克逊实验室的A/J JAX小鼠脑电图(EEG)中可以看到自发棘波放电(SWD),但来自Harlan的A/J Harlan小鼠没有此表现。
(6)行为学研究模型
与C57BL/6比较,A系小鼠呈现明显的焦虑样行为,较低的社会性和较少的运动活力。
(7)年龄相关的听力丧失模型和老年性耳聋模型
在4~5月龄开始发生听力丧失。原因是由于年龄相关听觉基因1突变。
(8) 呼吸系统疾病模型
香烟烟雾容易诱导肺气肿。实验诱导的哮喘可以诱导出类似于人哮喘的肺病理结构的改变。
(9) 补体C5基因缺乏的研究以及补体C5缺乏模型
补体C5是溶血性补体。A小鼠补体C5基因缺失。
(10)牙科模型研究
在比较12个亚系近交小鼠后,研究发现A小鼠是最容易发生氟斑牙,而129小鼠不敏感。

六、AKR系小鼠

特点:
(1)白血病:
自发性淋巴细胞性(胸腺的)白血病,白血病发生率60%~90%,发生时间:8月龄之后。含有ecotropic N-tropic murine leukemia virus。
(2)免疫学:
对绵羊红细胞低反应。用于Thy1.1(theta AKR)抗原的免疫学研究。从出生开始,具有毒血症,所有组织中表达亲嗜性逆转录病毒AKV。
(3)毛发育微细改变
原因是毛发育基因“hair interior defect”(hid)发生突变。鼠毛外观正常,但显微镜下可发现异常。
(4)肾上腺皮质脂剥夺
具有肾上腺皮质脂基因剥夺(ald),原因是sterol O-acyltransferase 1 (Soat1)基因突变。
(5)对饲料诱导动脉粥样硬化不敏感,对高脂饲料、高胆固醇饲料的反应性低。
七、KK小鼠
KK小鼠是一种轻度肥胖型2型糖尿病动物,1944年经近亲繁殖育成。
特点:
(1)为人类II型糖尿病模型,对双胍类降糖药敏感。
(2)1岁龄时多食、高血糖、高胰岛素血症、肥胖及肝脏对胰岛素的敏感性可自发恢复正常,但糖尿病KK小鼠生命常明显缩短。
(3)KK小鼠空腹胰高糖素水平升高,且不受葡萄糖抑制。
(4)组织学显示β细胞有脱颗粒和糖原浸润,随后出现胰岛肥大和肝脂肪化和脂肪组织增多。
八、FVB小鼠(白色)

特点及应用
(1) 生长发育强劲,活跃,生殖能力强
由于祖先是Swiss小鼠,从而保留了生长发育快、强壮、活跃和生殖能力强的特点,因而类似于Swiss小鼠,可以作为一般研究使用。
(2) 胚胎细胞核仁大
由于受精卵细胞核仁大有利于进行核仁注射、并且具有生长快繁育强的优点,.FVB小鼠广泛用于转基因鼠的母本。
(3) 对抗原刺激容易产生IgE
FVB小鼠很容易引发类似哮喘的呼吸道反应性,因此,呼吸道容易发生哮喘样改变。IgE形成具有高反应性,从而产生大量抗原特异性IgE。
(4) 对胰岛素诱导的低血糖反应性高。
(5) 视网膜退行性疾病
由于FVB小鼠Pde6brd1等位基因是纯合子,导致较早发生视网膜退化。
(6) 体温:应激时发生高热。
(7) 行为表现
FVB小鼠容易焦虑,适合作为某些行为研究模型,因而不适合作为研究行为的转基因的母本。
(8)肿瘤诱导
虽然FVB/N小鼠不容易自发产生肿瘤。化学诱导不容易发生乳头状瘤,但是一旦发生容易恶化为恶性肿瘤。化学诱导的结肠炎容易发展为直肠癌。
(9) 不适合用于胶原诱导关节炎模型
FVB小鼠虽然有MHC单体型H2q ,但对胶原诱导关节炎不敏感。
我国培育的近交系小鼠
01
中国一号(C1)小鼠
白色,繁殖力中等,肿瘤自发率低,1955年中国医学科学院实验医学研究所将昆明小鼠经近亲交配20代以上育成。
02
津白一号(TA1)小鼠
白色,于1955年从杂种白化小鼠和KM小鼠近亲交配育成,繁殖率中等,肿瘤发病率低。TAl为自发低乳腺癌系,易患慢性肺炎。
03
津白二号(TA2)小鼠
白色,于1963年从杂种白化小鼠和KM小鼠近亲交配育成,繁殖率中等。为自发高乳腺癌系,乳腺癌MA737的宿主,主要用于肿瘤学研究。
04
615小鼠
深褐色,1961年中国医学科学院输血及血液研究所将KM白化雌鼠和雄性C57BL近交系黑色小鼠杂交生产出第一代,然后再经近亲交配而育成。自发肿瘤发生率:低白血病、低乳腺癌、高肺腺癌。对白血病病毒和放射诱发白血病较敏感,广泛用于白血病研究和抗癌药物的筛选,同时还应用于肿瘤免疫、肿瘤机理研究。
05
SMMC/C和SMMC/B小鼠
1986年第二军医大学由KM小鼠近交培育而成,SMMC/C小鼠乳腺癌发病率高,对疟原虫敏感,SMMC/B小鼠肿瘤自发率低,对减压病敏感。
06
AMMS/1号小鼠(白化)
1974年军事医学科学院由KM小鼠近交育成,对炭疽弱毒株较敏感,对骨髓多向性造血干细胞测定比较规律。
07
NCPC/4
1992年由河北医学院、华北制药厂等以KM小鼠为原种,经26代以上严格近亲繁殖培育而成,繁殖能力强。对S180移植瘤的易感性较C57BL/6或NIH小鼠易感,并具有生长速度快的特点。易被白色念珠菌和微球菌感染。
08
SSB小鼠
1991年上海生物制品研究所选用昆明小鼠经近交34代后培育而成。该品系在微核、尿路上皮癌动物模型和F1代杂交诱生小鼠单克隆抗体等项实验中,具有一定的实验适应优于同类近交系,适宜作致癌、致畸、致突变实验,是理想的尿路上皮癌动物模型。
09
T739小鼠
土黄色,1987年由天津市医学科学研究所选用615公鼠和昆明母鼠近交46代后培育而成。肺腺癌LA795是该近交系小鼠的一种同基因型瘤株,所以该鼠是LA795的理想宿主。目前该系小鼠及其瘤株已广泛用于实验肿瘤学、 抗肿瘤药物的筛选和遗传学等方面的研究。它也是可移植性胸腹水型淋巴细胞白血病L845的同基因瘤株,已利用该模型进行多种抗癌药物的反应实验,比较敏感,效果良好。
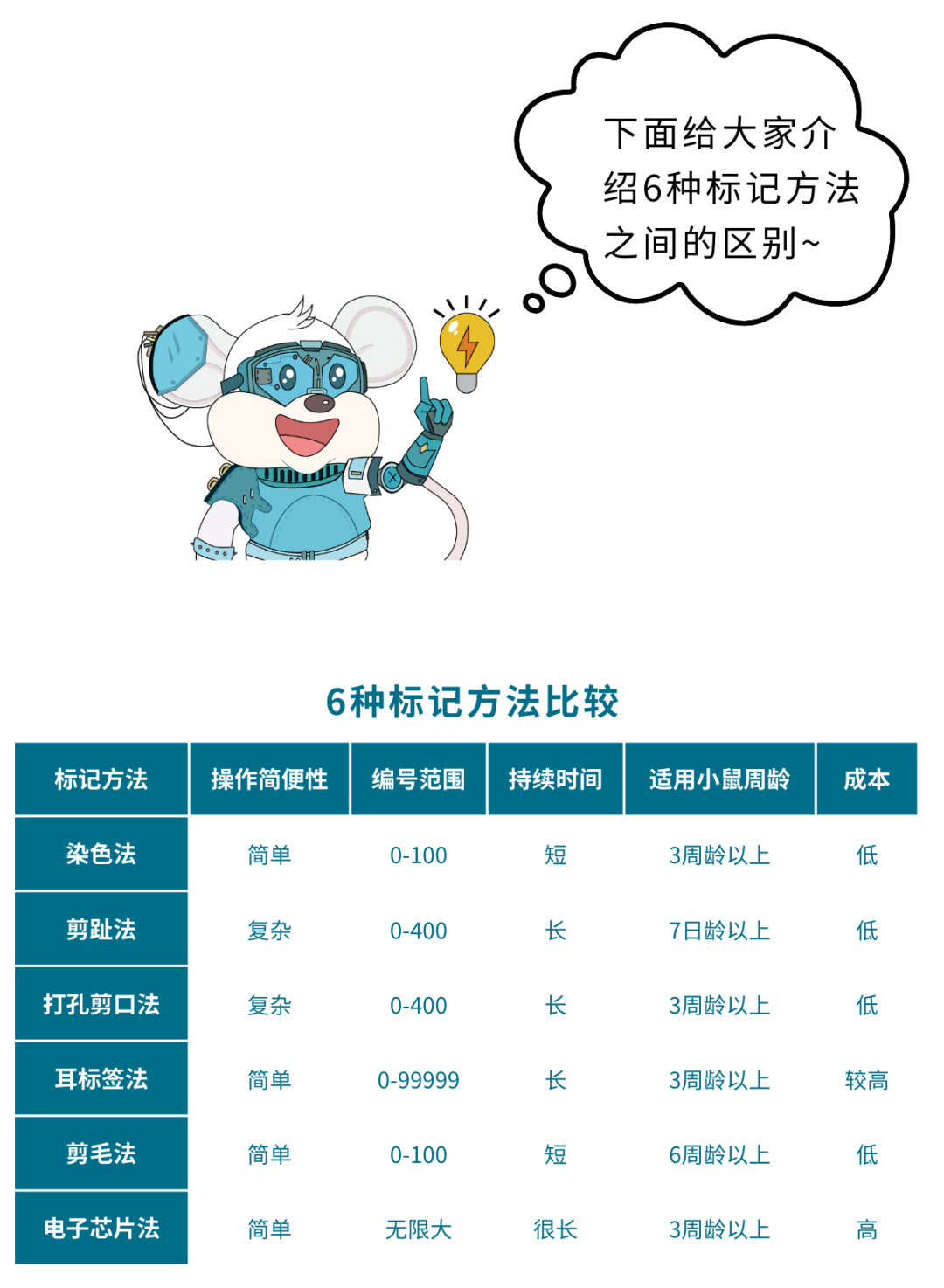
科研人当然不只有一只鼠啦,如何区分自己的上百只小鼠呢?为了防止混淆,我们需要清晰持久的编号或标记给小鼠编码,就像身份证号一样,可以明确区分大规模的鼠鼠。
身体染色法
用毛笔或棉签蘸取不同颜色的化学药品涂染在小鼠特定部位的毛发上,代表不同编号。常用的标记液有:3%~5%的苦味酸溶液(黄色);2%硝酸银溶液(咖啡色);0.5%的中性品红溶液(红色);煤焦油酒精溶液(黑色)。
编号的次序采用九分法原则,如图“先左后右、由前至后:左前肢上代表1号,腹部左侧为2号,左后肢上为3号,头顶为4号,腰背部为5号,尾基部为6号,右前肢上为7号,腹部右侧为8号,右后肢上为9号。小鼠同一部位一般最多可用两种不同颜色做标记,分别代表个位和十位。
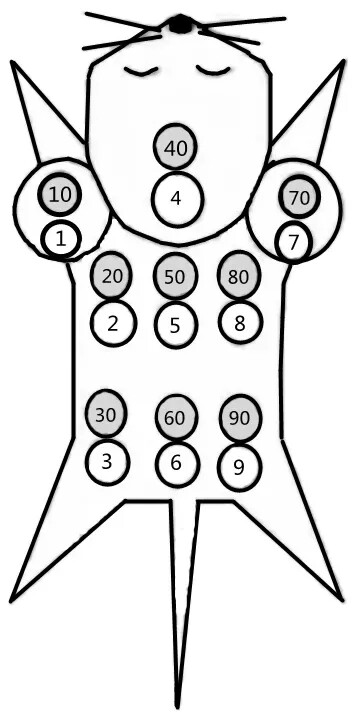
染色法的优点是操作简单,不伤害小鼠。缺点是由于小鼠体型小编号范围有限;只适合如ICR和BALB/C等毛色浅的品系,不适合毛色深的品系如C57BL/6小鼠;小鼠之间会相互扎堆嬉戏打闹,时间长了染色易褪色;苦味酸(2,4,6三硝基苯酚)是炸药的一种,属于国家限制使用的试剂;另外虽然这些染色剂属于低毒物质,但会通过小鼠理毛等行为进入体内,对实验结果的影响仍需要考虑。

尾巴染色法
用记号笔在小鼠尾巴上画出标记,适用于少量动物(<10只)和短周期的实验。如图:
尾巴染色法的优点是操作简单方便,易识别,不会给动物造成损伤及痛苦。缺点是时间太长易模糊,需再次标记。
耳标签法
用耳标钳等工具将刻有号码的耳标签订于小鼠耳部。耳标一般由塑料、铝或不锈钢等材质制成,编码范围为1~99999。

此法优点是编号范围足够大,操作方便快捷,编号清晰,持续时间长。缺点是小鼠周龄小时不适合打耳标签;耳标签容易被同笼小鼠弄掉,如果同笼内多只小鼠耳标签掉落,会造成小鼠编号无法确认给后续试验造成极大不便;耳标签不能重复使用,成本较高。

耳朵打孔剪口法
打孔法适用于成年小鼠或大鼠,先天性耳朵溃烂的某些品系和幼年鼠不太适用。此法优点编码范围较大。缺点一是操作不便,需要打孔器打孔和剪刀剪缺口;二是小鼠耳朵小,小孔或缺口等不易辨认;三是耳孔容易被同笼小鼠损坏,导致编码缺失或错误。
编号方式:使用打孔器和手术剪在对应编号位置将小鼠耳朵标记耳洞或者剪成缺口状,小鼠左耳可标记“个位数”,小鼠右耳可标记“十位数”,逢整百数不做标记。如图所示:
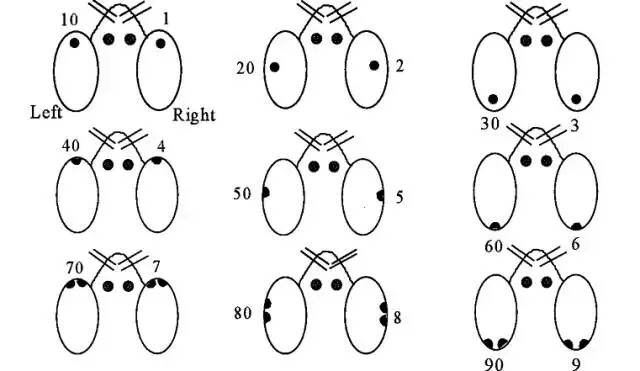
操作方式:单手抓取好小鼠颈后部皮肤,露出小鼠的耳朵,使小鼠腹部向下。不同的打孔器可能有不同的方法。

剪趾法
这种方法适用于出生5-7天,脚趾完整的幼仔(除如先天性脚趾畸形的某些特殊表型)。
编号方式:小鼠左下、右下脚趾依次标记为1~10号,左上、右上脚趾依次标记为20~90号。剪断相应的脚趾则代表对应的号码,逢整百数不做标记。(如:83号小鼠标记为剪断对应的80号脚趾和3号脚趾。100号小鼠标记为所有脚趾完好)。

操作方法:
1.操作前,操作人员手套表面浸泡或喷雾消毒。
2.左手抓取小仔鼠,腹部面向操作者,头部朝上,用左手食指和拇指固定住需要断趾的脚爪,用酒精棉球消毒脚趾。
3.用眼科剪(高压灭菌进入设施,使用前经75%酒精棉球消毒)剪去脚趾。
4.剪脚趾操作剪取的鼠爪量为鼠爪趾尖组织1~2mm,大约为整个脚趾的前端1/3,对应人的第一指节。尽量减少剪断的脚趾对小鼠运动的影响,也不能只断趾尖,以防伤口痊愈后辨别不清。
5.每取完一只小鼠,眼科剪需75%酒精擦拭消毒,以防小鼠交叉感染和样品污染。
6.如眼科剪疑似接触到污物,不得继续使用。
7.当天使用完毕后,眼科剪需要高压灭菌消毒。

剪毛法
在大鼠上有用剪刀按照染色法-九分法的原则剪去不同部位的适量毛发来做标记。此法同样可以用在小鼠身上。此法优点是操作简单,只用一把剪刀就可操作;对动物没有伤害,符合动物福利要求。缺点是编号范围有限,持续周期较短,不适合新生小鼠的标记。
针刺法
用七号或八号针头蘸取少量碳素墨水,在耳部、前后肢以及尾部等处刺入皮下,在受刺部位留有一黑色标记。该法适用于大小鼠、豚鼠等。在实验动物数量少的情况下,也可用于兔、犬等动物。
电子芯片法
动物电子芯片是一种利用无线电射频和微功耗集成电路技术,经过特殊封装工艺而制成的微电子芯片,信息量大,是世界上目前最先进的管理手段。晶片是一种小型已写入鉴定序号的计算机芯片,装在一个很小的表面光滑且不会引起动物排斥的玻璃体内,注射前将玻璃体放在皮下注射器针头里然后注射到动物皮下。动物一经注射晶片,就具有唯一的序列号,可以一直被识别,不会丢失和改变。注射芯片后不会影响动物的饮食饮水以及生长繁育。读取编号时用电子阅读器在笼盒外部接近动物到一定距离时即可读取到动物的编号。整个读取过程不用抓取动物,对动物没有刺激。
该方法的特点是;编号范围广;标签在体内,不易破坏;使用寿命长,可达20多年;可循环使用;读取简单,不用每次抓取动物进行读取,对动物应激小。缺点是不适合短期实验;成本很高;对于皮下接种的一些实验电子芯片的存在可能会有影响。
在实验猴或树鼩的研究中有用电子芯片对其进行标记。实验猴或树鼩由于行动灵活,不易抓取等特点常规的标记方法不太适用,所以研究出了电子芯片标记的方法和仪器。
电子芯片同样也可以应用到大小鼠的标记中。对于长期的或者日常实验可以自动化、高通量化的小鼠实验,也可以考虑用这种标记方法。
繁殖担任着物种延续的崇高使命,帮助实验小鼠繁殖,也是科研人必不可少的工作之一,怎么配繁?
配繁时期及小鼠选择
一、小鼠的性成熟
一般雌鼠性成熟期为4-6周,雄鼠为6-8周。性成熟的表现是生殖器官发育完全,出现第二性征。雌鼠具有产卵子的能力,雄鼠具有产精子的能力。
雌鼠性成熟标志:雌鼠阴道口张开,出现求偶周期,有交配的欲望,愿意接近雄鼠。
雄鼠性成熟标志:雄鼠睾丸自腹腔落入阴囊,且能产生精子。对于雄鼠,从6周龄开始可先将其与野生型雌鼠合笼,进行交配前的训练,证明雄鼠没有问题,到8周龄时,再对雄鼠进行真正的使用。
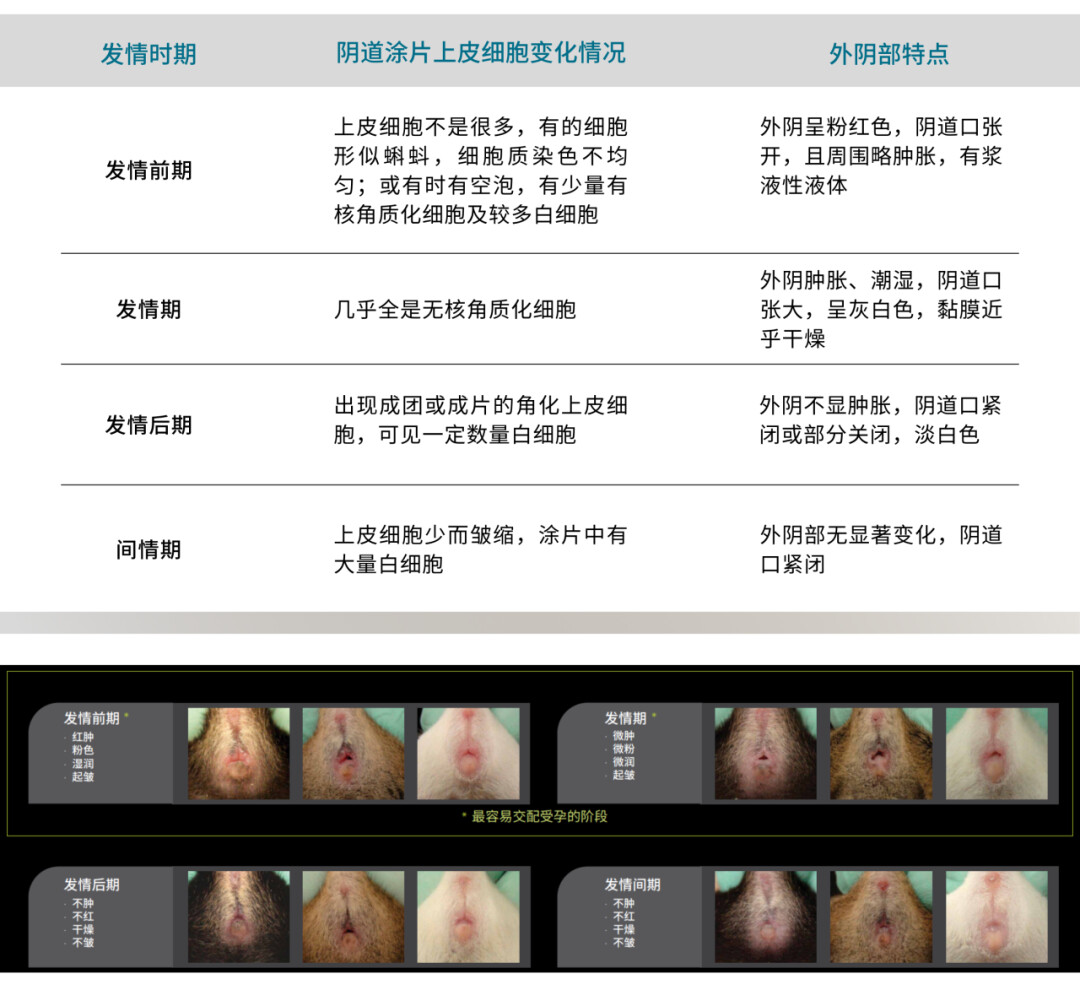
二、小鼠的性周期
雌鼠全年多次发情,性周期为4-5天,一般可分为发情前期,发情期,发情后期和休情期。一般在发情后2-3个小时即可排卵,排卵期3-4天,但在排卵期仅有数小时允许雄鼠交配。产后12-14小时内会再有一次发情。自此,由于雌鼠需给仔鼠哺乳,性周期会暂时停止。空窗独自生产的雌鼠,在仔鼠离乳后2-6天可出现下一个性周期(考虑到动物福利,不建议母鼠连续性繁殖哦)。

三、小鼠的mating
小鼠一般选择在22:00——01:00交配,可以一雄一雌或一雄多雌合笼。根据合笼时间,可分为连续合笼和间断合笼,连续合笼即雌鼠和雄鼠一直处于一个繁殖笼中,间断合笼指一只雄鼠和雌鼠相处一天,次日再换其它的雌鼠进行合笼。一般来说,长期合笼可以提高怀孕率,因为小鼠更喜欢熟悉的配偶。但一夫多妻和间断合笼效果也不一定差,因为有时雄鼠不太喜欢“包办婚姻”。

鼠鼠的妊娠和哺乳
一、我的小鼠怀上了吗
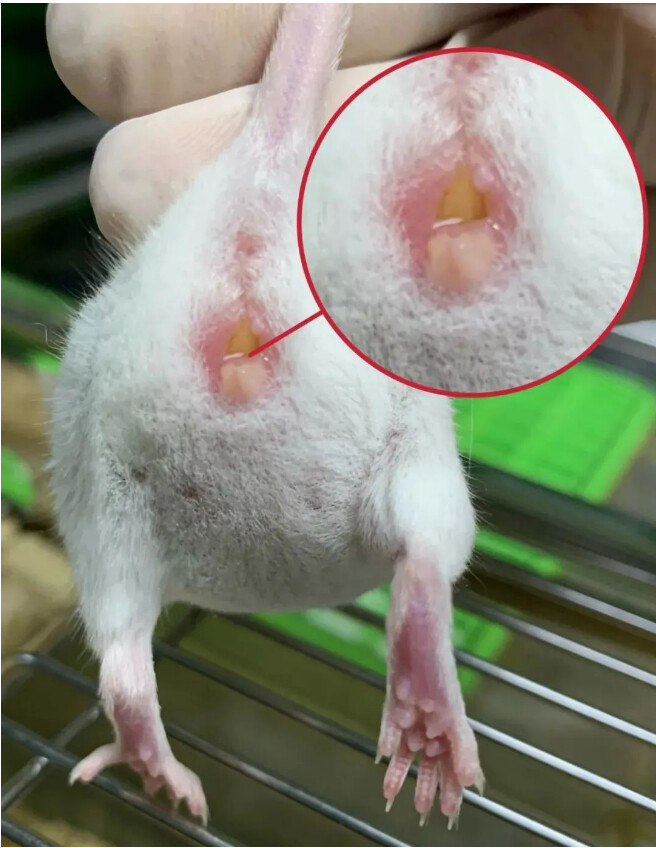
小鼠一旦干过羞羞的事,就会留下犯罪证据。两鼠交配后4~5min,在雌鼠的阴道口会形成一种白色浆液性物质堵塞在阴道至子宫颈的腔内,称阴道栓(vaginalplug)形,这是雄鼠的精液、雌鼠的阴道分泌物和阴道上皮混合遇空气后变硬的结果,可防止精子倒流,提高受孕率。阴道栓常视为mating成功的标志,看见栓的中午12点,记为胚胎期0.5天。阴道栓在交配后12-24小时会自动脱落。

小鼠妊娠期为19-21天,怀孕后10-12天可触摸到胎儿,轻轻感觉到下腹部有“小圆球”,但是触摸时要注意跟粪便区分,以及要控制手指的力度,若太用力会导致胚胎溶解,妊娠会终止。

二、小鼠的分娩
小鼠的分娩可昼夜进行,但以晚间为多,整个分娩过程约一小时。母鼠产前一般是表现不安状态,常常不停地整理产窝。约四分钟产仔一只,一分钟后胎盘产出,母鼠将胎盘嚼食。母鼠产后12-24小时会再次发情,此时就交配可造成产后妊娠(边哺乳边怀孕),这种交配方式,妊娠期大概率比一般妊娠期长,可达30天左右(考虑到动物福利,不建议这样做哦)。


小鼠每胎产仔6-15只,产仔数取决于品系、胎次、饲养条件、营养条件等,第2-6胎产仔数较多,一般7胎后产仔数逐渐下降。
三、哺乳
哺乳期18-21天,小鼠带仔数一般为8-10只,具体根据母鼠营养状况、体质状况、生产能力等因素的不同会有变化。哺乳仔数太多可能导致部分仔鼠发育不良,因此可提前准备代乳母小鼠帮助哺育后代。
四、生育寿命
小鼠性活动可维持1年左右,随着年龄的增长,小鼠变的嗜睡,不爱活动,繁育能力逐渐下降,雌鼠会出现难产,食仔,幼仔畸形等。雌鼠的生育寿命6-12个月,雄鼠的生育寿命12-14个月,超过年龄的小鼠应淘汰。
小鼠属脊椎动物门,哺乳纲,啮齿目,鼠科,小鼠属动物。小鼠的世代短,成熟早,繁殖很强,属全年、多发情动物,一年可产仔胎数6-10胎,每胎产仔数8-15只,且具有体型小、易于管理等特点。在进化上,小鼠与人类的距离较近,其解剖学结构、发育过程、生化代谢途径都与人接近,为人类疾病的克隆提供了极佳的实验材料。作为经典的医学实验动物,小鼠已经对医学诸多领域的研究作出了卓越贡献。

从宠物到实验动物
大约6000万年前,鼠属出现在地球上,经过漫长的繁殖和进化,出现诸多分支,比如褐家鼠、仓鼠、田鼠等。
早在3000年之前人类已经注意到小鼠性状的遗传。
在公元前1100年中国已经使用花老鼠(spotted mice)一词。

1664年,Robert Hooke用小鼠进行大气研究,标志着小鼠开始作为实验动物来使用。
直至19世纪,宠物爱好者开始选择性的饲养和驯化家鼠,从而逐渐出现了一些具有各种可爱颜色皮毛的宠物鼠。
1865年,奥地利业余生物学家(主业是修道士)孟德尔通过豌豆实验,得出了著名的孟德尔定律。

1897年,生物学家William Haacke发现白鼠和杂色鼠杂交会产生重组表型。

1900年,尘封35年的孟德尔遗传理论被欧洲的科学家“重新发现”,在科学界掀起了在不同物种中验证孟德尔遗传定律的风潮,小鼠性状的不连续变异非常类似于孟德尔在豌豆中的遗传发现。
William Ernest Castle和Lucien Cuénot的工作证明了孟德尔遗传定律在动物中也是适用的,从此开启了哺乳动物遗传学研究的时代。
现代近交系小鼠开发的鼻祖——Castle和Lathrop
20世纪初期,小学老师Abbie E.C.Lathrop决定下海经商,她搬到马萨诸塞州的格兰比农场开始了家禽养殖的生意,1900年家禽生意失败后,她开始在农场中饲养并出售来自世界各地不同品种的小鼠。

Lathrop的这些老鼠引起了遗传学家William E. Castle的注意。
1902年,哈佛大学的William Ernest Castle通过观察的宠物鼠毛色,以验证孟德尔遗传定律的正确性。从此启动了当代小鼠基因研究。

当时养老鼠的Lathrop已经小有名气,并且她对小鼠的繁殖有着非常详细的计划并且饲养记录也是非常完备的。
在有严格记录下饲养的小鼠对研究小鼠的遗传是非常有帮助的,所以慢慢地各个科研机构的研究者们开始向她订购老鼠,用于科学研究。
1905年,法国遗传学家Lucien Claude Cuéno对黄白相间的杂色鼠进行研究,现两个携带黄色皮毛基因的宠物鼠之间交配有纯合致死现象。

1909年遗传学家Clarence Cook Little培育出了第一个现代实验鼠株系——DBA小鼠(毛色为淡棕色),这意味着现代实验室小鼠的诞生。

1908年Lathrop发现她的一些老鼠出现了不寻常的皮肤病变。她将样本寄给了在宾夕法尼亚大学的实验病理学家Leo Loeb,经过他研究,确定了这些病变的为恶性病变。

1913年-1919年Lathrop与Leo Loeb在著名期刊上先后发表了10篇期刊论文,如“实验医学杂志”和“癌症研究杂志”。
小鼠动物实验正式拉开了序幕。
C57BL/6J小鼠

1918年,一战服役结束后,C.C.Little来到了冷泉港。
1921年C.C.Little用Lathrop的小鼠近亲培育数个近交系,使编号为57的雌鼠与52号雄鼠交配,培育成C57。其中毛色固定为黑色的培育成C57BL,BL是英文Black(黑色)的缩写。
1937年,Little将维持的C57BL父系进行分离,将第6组亚系定名为C57BL/6,而将第10组亚系定名为C57BL/10。
1947年,美国Jackson Laboratory从Little处引进C57BL/6进行培育,命名为C57BL/6J,6J中的J指的是Jackson Laboratory。
1951年,美国国立卫生研究院(National Institute of Health,NIH)从Jackson Laboratory引入了C57BL/6J第32代并形成了亚系C57BL/6N,6N中的N指的是NIH。
不同单位近交培育的C57BL/6N在其后追加标记,比如1991年,Taconic Farms近交培育C57BL/6N并命名为C57BL/6NTac。

BALB/c小鼠

1913年纽约纪念医院的Halsey J.Bagg买了一些小白鼠用于研究。Bagg把自己名字里的Bagg和白化英文Albino糅合在一起,取名BALB。

1920-1935年期间,Bagg系统地将这些小白鼠的后代同胞兄弟姐妹进行了近交26代。
1935年,George Davis Snell(美国著名遗传学家,免疫遗传学奠基人,因发现遗传因素决定个体间移植组织或器官存活状况并最早提出了H-抗原概念获得1980年诺贝尔生理学与医学奖)将他们引种到杰克逊实验室。他在BALB名字后加了个小写的c字母,表示毛色隐性上位作用。Snell慷慨地把BALB/c分享给同事J. Paul Scott和美国国立卫生研究院(NIH)进行维护。

1947年,杰克逊实验室发生异常火灾,Snell失去了所有小鼠品系。他想到曾经送给Scott的一些BALB/c品系。Scott将他养的第41代BALB/c还给了Snell(现在称为BALB/cJ,截至2006年12月14日,它们总代数为221代)。
1985年我国从美国NIH引进BALB/c第180代。

裸鼠
20世纪60年代初,是英国格拉斯哥鲁契尔医院病毒实验室的Grist博士最先在其饲养的白化种群小鼠(非近交系)中偶然发现有个别天然无毛的小鼠。
1966年,爱丁堡动物遗传研究所的Flanagan从Grist处寻得3只裸鼠,通过遗传和生物学特性分析,确定出了无毛突变基因杂合子母代,利用这对杂合子和它们的后代建立了裸鼠群,用“nu”表示该突变基因,并命名为Nude小鼠。
至此,无毛“裸鼠”终于有了自己的名字。Flanagan也因此项成果在Genet Res发表了一篇名为《“Nude”, a new hairless gene with pleiotropic effects in the mouse》的文章,开启了免疫缺陷实验动物研究的新篇章。

1968年,苏格兰斯特拉斯克莱德大学生物学系的Pantelouris从爱丁堡动物遗传研究所得到了裸鼠,并在解剖过程中意外地发现裸鼠胸腺发育不全。而早在上世纪40年代,为开展细胞免疫学的研究,通常要使用药物来抑制小鼠T细胞的产生或通过外科手术摘除胸腺。无胸腺的“裸鼠”,无疑成为了可替代胸腺切除小鼠的一种天然免疫缺陷动物模型,这一裸露表型的重要发现,使Pantelouris的工作引起了科学界的广泛关注。
1985查尔斯河实验室通过BALB/cABom-nu和BALB/cAnNCrj-nu进行杂交和回交从而获得CAnN.Cg-Foxn1nu/Crl的纯种雌性,自此产生了常用的自交系小鼠BALB/c-nu。
SCID小鼠

SCID小鼠是继裸鼠出现之后,人类发现的又一种十分有价值的免疫缺陷动物,由位于小鼠16号染色体上的Prkdc (Protein kinase, DNA activated, catalytic polypeptide,又名scid)基因突变而来。
1980年Bosma从Fox Chase癌症中心饲养的C.B-17/lcr小鼠群中发现scid的单个隐性基因发生突变,C.B-17小鼠是BALB小鼠的同源近交系,该小鼠除了携带来自C57BL/Ka小鼠的免疫球蛋白重链IG-1b等位基因与BALB小鼠不同外,两品系小鼠其余基因完全相同。
1983年美国学者Bosma等发现C.B-17纯系小鼠16号染色体上的重症联合免疫缺陷(SCID)基因发生隐形突变后,小鼠缺乏功能成熟的T和B淋巴细胞,而称为SCID鼠。
1992年,Jax实验室用非肥胖糖尿病小鼠NOD/Lt与SCID小鼠杂交得到了NOD SCID小鼠,这种小鼠除淋巴细胞缺陷外,巨噬细胞功能、补体依赖溶血活性、NK细胞活性均降低,肿瘤接种成功率明显提升。但NOD SCID小鼠平均8.5个月发生胸腺瘤,不宜做长期模型。
2005年,Jackson Lab用白细胞介素IL-2Ry链等位基因突变鼠与NOD SCID小鼠回交形成NSG小鼠,NSG小鼠是SCID突变和IL-2Ry等位基因突变的双重纯合子。NSG小鼠能存活16个月,是目前构建PDX(人源性组织异种移植)模型的最佳小鼠。
“定制小鼠”时代与动物福利
20世纪70年代末80年代初,分子生物学的兴起将小鼠研究带入了基因修饰的新时代。
1980年,耶鲁大学的Jon Gordon和Frank Ruddle通过原核注射受精卵创造了第一只转基因小鼠。
1981年,剑桥大学遗传学家Martin Evans等创造了第一个小鼠多能干细胞系,为转基因小鼠做好铺垫。美国犹他大学分子生物学家Mario Capecchi和Oliver Smithies在此基础上制出第一批基因敲除小鼠,和Martin Evans共享了2001年拉斯克生物医学研究奖。

2013年,31岁的美国麻省理工学院教授张锋在《Science》上发表论文,宣布用CRISPR-Cas9系统首次实现对小鼠基因的编辑,并确认它能在几周内建立起小鼠的疾病模型。基因魔剪横空出世,指哪打哪,癌症、艾滋病、镰刀状红细胞贫血症……经典的人类疾病通过改写基因在小鼠身上重现,经受各类药物和技术的检验。经过遗传改造的小鼠也越来越适应科研人员的不同研究需要,会有更多的小鼠沿着它们祖祖辈辈的道路献身科学。

随着近年来生命科学技术蓬勃发展,医学研究对动物实验的依赖越来越得到加强。医学的前进与发展,每一步都留下了实验动物的足迹。出于对实验动物的基本权利的维护,倡导人与自然和谐共处与发展,更加为人类的健康服务,下面介绍动物实验中的一些自由原则及3R原则。

3R原则并不是要求不用动物,而是更希望科研人员优化设计好实验,改进实验设计和规范实验操作。动物中心保障动物的质量、标准化饲养环境和方式,保证实验数据的可靠性、准确性和科学性。
动物和人一样,有大脑思维、有喜怒哀乐、有疼痛感、有恐惧感,大自然给予它们同等的生存权利。在实验动物的使用和研究中坚持3R原则,要善待活体动物,减少痛苦和死亡率。例如,给予动物舒适的居住环境、足够营养的饲料、清洁的饮用水;给予必要的镇静剂、麻醉剂;温和保定、善良抚慰、减少应激、实施安乐死等。